
Picture a watchdog so vigilant it never lets its charge out of sight, holding tight to prevent any escape. That’s KEAP1, a protein that grips onto NRF2 with remarkable tenacity. But when cellular stress strikes, this molecular partnership transforms into one of biology’s most elegant release mechanisms.
What is the NRF2-KEAP1 system
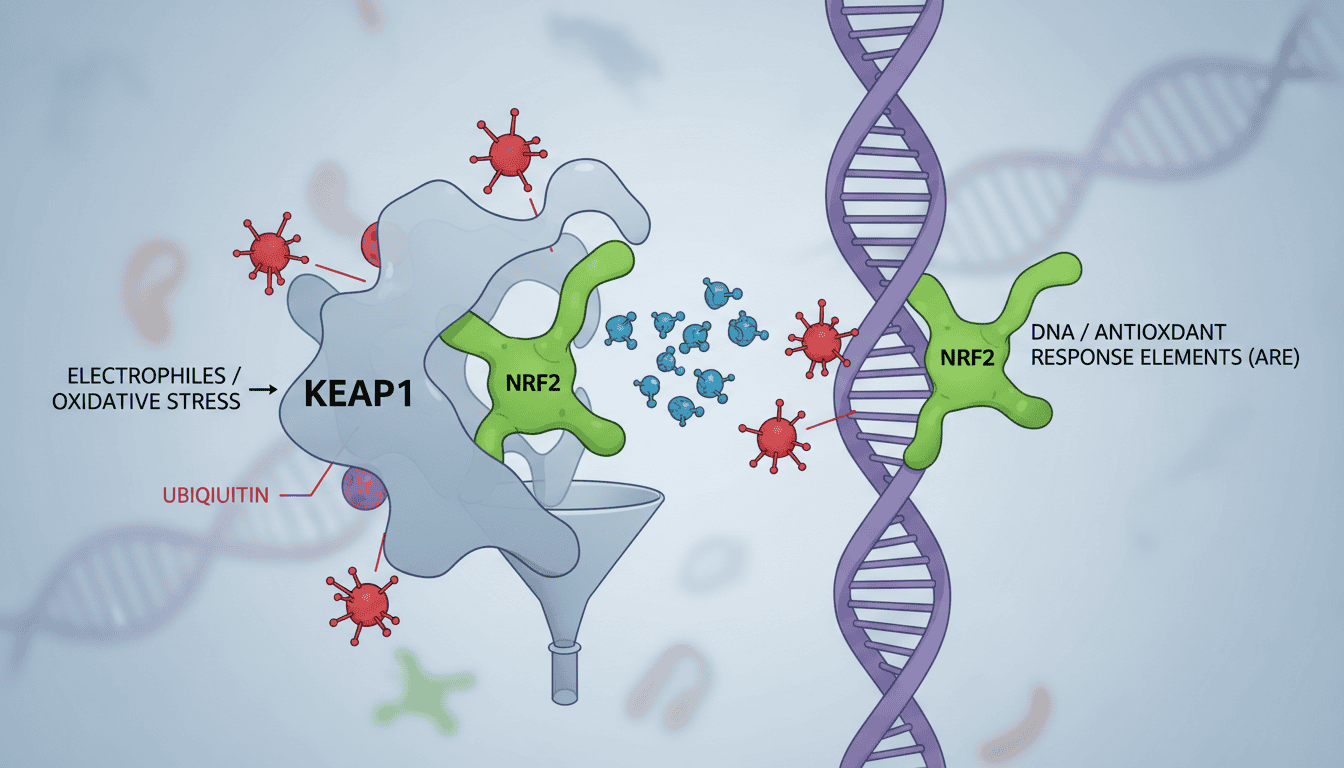
NRF2 and KEAP1 form a control system that governs how cells respond to oxidative stress. Think of KEAP1 as a molecular handcuff. Under normal conditions, it binds tightly to NRF2, keeping this transcription factor locked away in the cell’s cytoplasm.
KEAP1 doesn’t just hold NRF2 prisoner. It actively marks it for destruction by recruiting enzymes that tag NRF2 with ubiquitin molecules. These tags act like a “demolish this” sign, sending NRF2 to the proteasome where it gets broken down into amino acids. The result? NRF2 levels stay low, and the cell’s antioxidant defences remain quiet.
This system evolved as a hair-trigger response mechanism. When oxidative stress appears, certain amino acids in KEAP1 get chemically modified. These modifications weaken KEAP1’s grip on NRF2, allowing the transcription factor to slip away and rush to the nucleus. Once there, NRF2 switches on dozens of protective genes simultaneously.
What the research shows
Scientists have mapped this interaction down to individual amino acids. KEAP1 contains highly reactive cysteine residues that act as molecular sensors. When reactive oxygen species or electrophilic compounds encounter these cysteines, they form covalent bonds that alter KEAP1’s shape.
The most studied of these sensors is cysteine-151. Researchers found that modifying this single amino acid can trigger NRF2 release. But KEAP1 contains multiple cysteine sensors, each responding to different types of cellular stress. This gives the system remarkable sensitivity to various threats.
Laboratory experiments reveal the speed of this response. Within minutes of stress exposure, NRF2 levels in the nucleus jump dramatically. The transcription factor then binds to antioxidant response elements in gene promoters, ramping up production of enzymes like glutathione S-transferases, NAD(P)H quinone oxidoreductase, and haem oxygenase-1.
Structural studies using X-ray crystallography show exactly how KEAP1 grips NRF2. The interaction involves two binding sites on NRF2 that fit into KEAP1 like a key in a lock. When KEAP1’s cysteine sensors get modified, this lock mechanism fails, freeing NRF2 to activate cellular defences.
Why cells need this system
The tight control that KEAP1 exerts over NRF2 prevents unnecessary activation of antioxidant genes. Running these protective systems constantly would waste cellular energy and resources. Cells need to respond to genuine threats, not false alarms.
This system also provides proportional responses. Mild oxidative stress triggers modest NRF2 activation, while severe stress overwhelms more of KEAP1’s sensors, leading to stronger defensive responses. The multiple cysteine sensors act like a volume control, allowing cells to match their response to the threat level.
The rapid degradation of NRF2 under normal conditions ensures the system can reset quickly. Once the stress passes and KEAP1’s sensors return to normal, newly made KEAP1 proteins quickly capture any remaining NRF2 molecules. This prevents the antioxidant response from lingering longer than necessary.
Evolution preserved this mechanism because it balances protection with efficiency. Cells that couldn’t mount antioxidant responses died from oxidative damage. But cells that ran these expensive protective systems all the time got outcompeted by more efficient neighbours.
What affects NRF2-KEAP1 interaction
Age alters this system in multiple ways. Older cells often show decreased ability to activate NRF2, partly because KEAP1’s cysteine sensors become less responsive to modification. The proteasome machinery that normally degrades NRF2 also becomes less efficient with age, creating a complex shift in the system’s dynamics.
Exercise triggers NRF2 activation through multiple pathways. The oxidative stress generated during physical activity modifies KEAP1’s sensors, while mechanical stress on cells activates additional signalling cascades that converge on this system. Regular exercise appears to maintain the sensitivity of KEAP1’s stress sensors.
Certain foods contain compounds that interact directly with KEAP1. Sulforaphane from broccoli, curcumin from turmeric, and various polyphenols can modify KEAP1’s cysteine residues, mimicking the effects of endogenous stress signals. However, the bioavailability and tissue distribution of these compounds varies significantly.
Environmental toxins, air pollution, and chronic inflammation all affect this pathway. Some toxins damage KEAP1’s sensors, making them hyper-responsive. Others interfere with NRF2’s ability to bind DNA once it reaches the nucleus. The system that evolved to protect cells can become a target for environmental damage.
What remains unknown
Researchers still puzzle over why some tissues show stronger NRF2 responses than others. The liver and kidneys activate NRF2 powerfully, while the brain shows more modest responses. Whether this reflects differences in KEAP1 expression, alternative regulatory mechanisms, or tissue-specific factors remains unclear.
The relationship between NRF2 activation and cellular ageing continues to generate debate. Some studies suggest that maintaining NRF2 activity supports healthy ageing, while others indicate that excessive activation might promote certain disease processes. The optimal level and duration of NRF2 activation for different life stages isn’t well understood.
Scientists are also investigating whether KEAP1-independent pathways for controlling NRF2 play significant roles in human health. Alternative mechanisms involving different proteins might fine-tune NRF2 activity in ways that current research hasn’t fully mapped.
The precision of cellular stress sensing through the NRF2-KEAP1 system reveals how evolution crafted molecular machines of remarkable sophistication. This partnership between a vigilant guardian and a powerful responder shows that even at the cellular level, protection requires both sensitivity and restraint. As researchers continue mapping these molecular interactions, they uncover not just how cells defend themselves, but how the fundamental logic of biological systems balances competing demands for survival and efficiency.
Matt Elliott is the editor of Redox News Today, an independent publication covering peer-reviewed research on cellular health, redox signalling, and related biomedical science.




